Protein A antibody-binding domains fold independently
Circular dichroism and fluorescence across all five domains of SpA — in isolation and in full-length SpA-N — reveal a steep N-to-C terminal stability gradient spanning a wide free-energy range.
THE CHALLENGE
Five tandem domains — coupled, or independent?
Staphylococcal Protein A's IgG-binding region (SpA-N) is a tandem repeat of five small, homologous domains. A natural question for any engineered or therapeutic construct: do the domains stabilize one another, or does each fold on its own? And if they fold independently, how much does their stability actually vary?
Bulk measurements on the full-length protein average over all five domains — obscuring exactly the per-domain differences that matter for stability engineering.
WHAT WE DID
Measure every domain, in isolation and in context
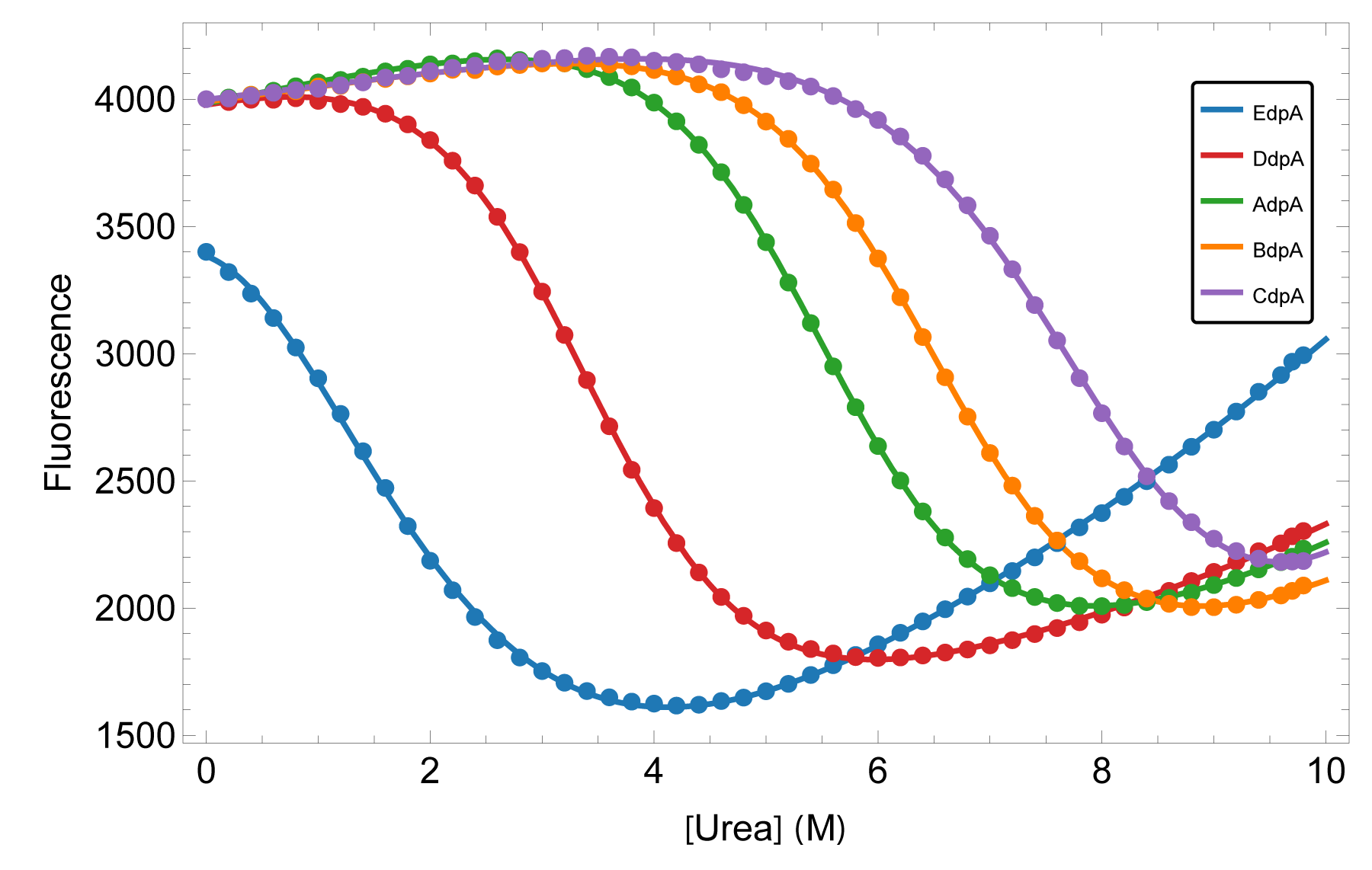
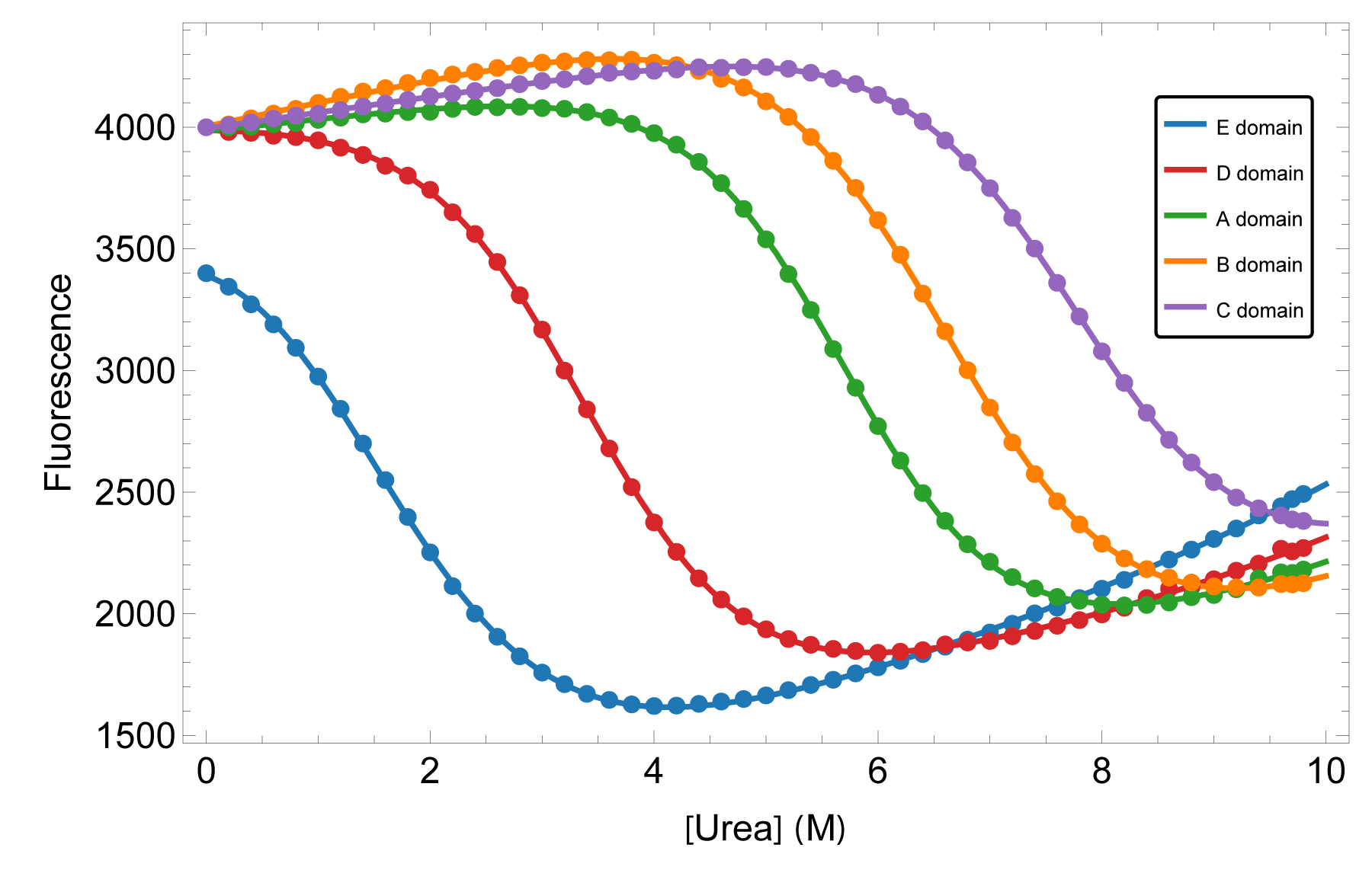
We characterized the equilibrium stability of all five domains both as isolated constructs and within the full-length protein, using circular dichroism and fluorescence to follow folding. Comparing isolated-domain behavior to the same domain in context tests directly whether neighbors perturb one another — and a global thermodynamic treatment turns the curves into per-domain free energies.
That side-by-side design is what separates “independent” from “coupled” rather than assuming either.
WHAT IT REVEALED
Independent domains, a wide stability range
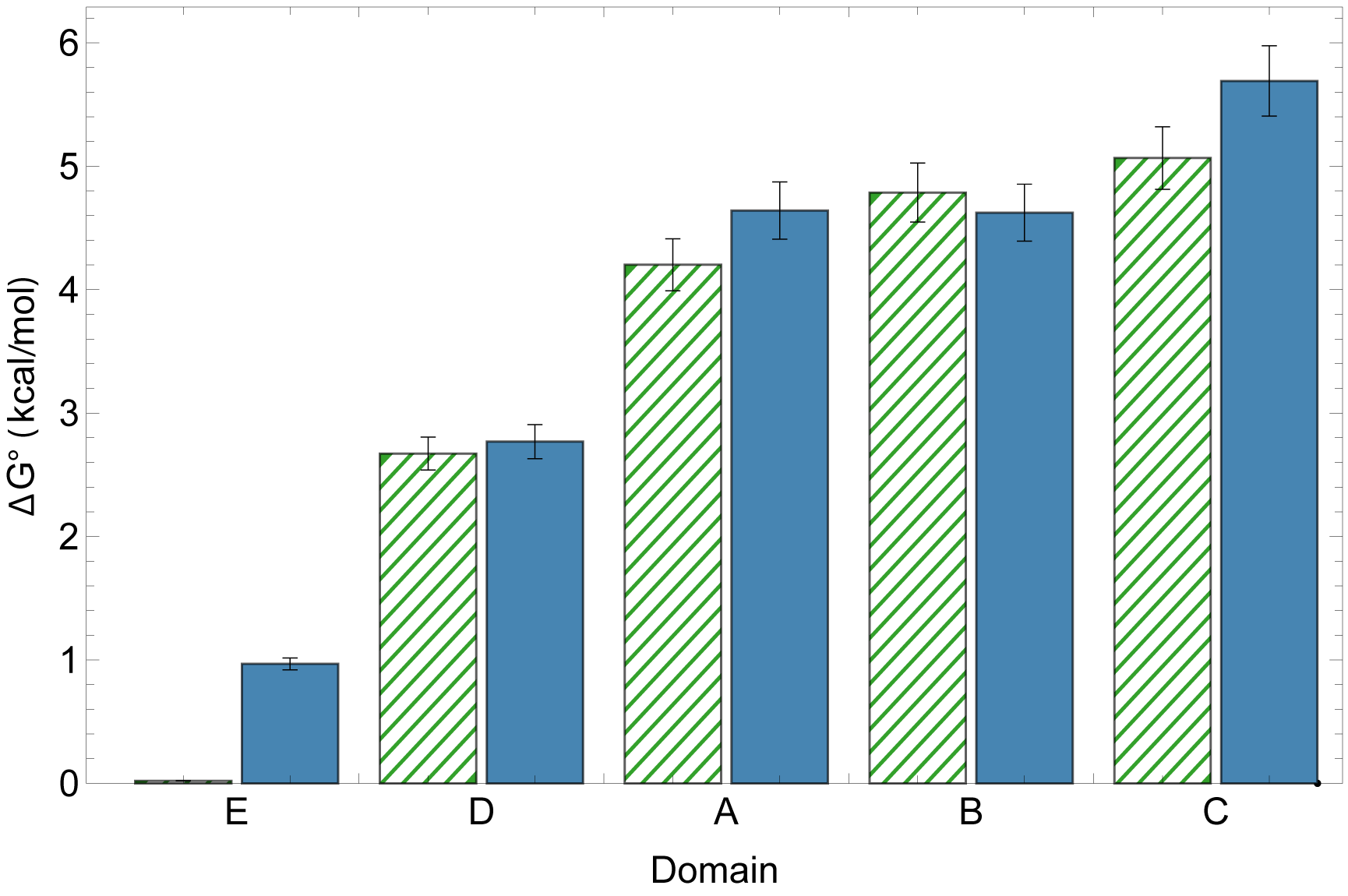
The domains fold independently — their behavior in full-length SpA-N matches their behavior in isolation — yet their stabilities are far from uniform. A steep gradient runs from the N-terminal to the C-terminal domain, so the construct carries both its most and least stable elements in a defined order. For engineering and formulation, that means the weakest link is identifiable, not hidden in an average.
A possible role for the gradient in secretion
The gradient may not be incidental. SpA is secreted N-terminus first through the Sec translocon, and we hypothesize the stability gradient facilitates efficient co-translational secretion: a weakly folded N-terminal E domain (ΔG° ≈ 1 kcal/mol) presents far less of a kinetic barrier to threading than a stable domain would, while the more stable C-terminal domains benefit from remaining in the cytoplasm longer before translocation.
Independence plus a wide stability range means each domain can be tuned on its own — once you know which one sets the limit.
Conformational change coupled to ligand binding
A 10-state model resolving conformational selection and induced fit — operating together near the apparent Kd.
Read case study →Exposing hidden high-affinity states
A 6-state model: the highest-affinity state is present all along, hidden by its low population.
Read case study →Have a system like this?
Tell us about the protein, the data, and the question. We'll tell you whether a thermodynamic model can answer it.
Contact Us→